
金乌贼成体副缠卵腺的超显微结构
金乌贼成体副缠卵腺的超显微结构
刘长琳 赵法箴 边 力 葛建龙 谭 杰 陈四清①
(1. 农业部海洋渔业可持续发展重点实验室 中国水产科学研究院黄海水产研究所 青岛 266071;2. 青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071)
头足类生殖系统主要由性腺、生殖导管和附属腺组成(林东明等, 2013),是头足类的重要系统之一,不仅与生殖活动密切相关,还是头足类分类的重要依据(刘必林等, 2010)。因此,开展头足类生殖系统研究,不仅是生物学研究的基础,也是繁殖生物学研究的重点。副缠卵腺与缠卵腺统称为附属腺,是乌贼目(Sepiida)中雌性亲体生殖系统的重要组成部分,与生殖活动密切相关(王春琳等, 2010)。关于头足类副缠卵腺的研究报道相对较少,国外仅见福氏枪乌贼(Loligo forbesi)副缠卵腺组织学(Lum-Kong et al,1992b)、皮氏枪乌贼(Loligo pealei) (Bloodgood, 1977)和虎斑乌贼(Sepia pharaonis)(Nair et al, 2005)副缠卵腺的超微结构,以及副缠卵腺共生菌(Lum-Kong,1992a; Barbieri et al, 1997; Pichon et al, 2005)等方面的研究,国内仅见曼氏无针乌贼(Sepiella maindroni)副缠卵腺组织学和超微结构研究(蒋霞敏等, 2008;王春琳等, 2010),目前,未见金乌贼副缠卵腺的相关研究报道。本研究揭示了金乌贼(Sepia esculenta)副缠卵腺的超显微结构,并探讨了副缠卵腺的功能和作用,旨在揭示金乌贼的繁殖策略,丰富头足类繁殖生物学内容,进而为金乌贼繁育技术的完善提供参考资料。
1 材料与方法
1.1 材料
雌性金乌贼亲体于2016年6~7月在青岛黄岛海域(120°20′~120°38′E、35°74′~35°92′N)采用地笼网捕捞获得,均处于产卵盛期,平均体重为(795.6±18.4) g,平均胴长为(19.4±1.8) cm (n=10),活体塑料袋充氧运输至青岛金沙滩水产开发有限公司金乌贼实验基地。
1.2 方法
1.2.1 取样 将金乌贼称重后,沿腹中线剪开,取副缠卵腺,测量称重后,切成小块。
1.2.2 石蜡切片样品的制备 将0.5 cm×0.5 cm×0.5 cm的副缠卵腺小块用Bouin氏液固定,24 h后转入70%的乙醇中保存。梯度乙醇脱水、二甲苯透明、浸蜡、石蜡包埋,KD-2508型轮转式切片机连续切片,切片厚度为5~6 μm,HE染色,中性树胶封片,晾干后于Nikon 80i 正置显微镜下进行观察拍照。
1.2.3 透射电镜样品的制备 将体积为(0.5~1) mm3副缠卵腺小块于2.5%戊二醛中固定4 h,PBS缓冲液冲洗,1%锇酸4℃后固定2 h。PBS缓冲液冲洗后,乙醇系列梯度脱水,Epon812环氧树脂包埋。UltracutE超薄切片机半薄切片,甲苯胺蓝染色,半薄定位,醋酸双氧铀硝酸铅染色。采用JEOL公司型号为JEM-1200EX的透射电镜进行观察。
2 结果
2.1 副缠卵腺的解剖结构
金乌贼副缠卵腺1对,位于缠卵腺的前端,开口于外套腔,颜色为白色并夹杂着橙黄色,未见成熟晚期变红趋势。副缠卵腺形状不规则,呈耳状,沿腹中线左右对称,上部外缘中间向下凹陷,下部外缘位于缠卵腺两侧,且中部向下延伸,使得下端呈‘M’型与缠卵腺紧密连接。副缠卵腺的重量为1.32~4.10 g,宽度为 3.70~5.36 cm,高度为 2.64~3.27 cm (n=10)(图 1)。
2.2 副缠卵腺的显微结构
金乌贼副缠卵腺结构主要包括腺壁、腺体小管和结缔组织3部分,其中,腺壁包括由单层柱状上皮细胞组成的外膜层和平滑肌组成的肌肉层,外膜层厚度约为25~40 μm,肌肉层不同部位厚度存在较大差异,最厚可达0.3 mm (图2A)。腺体小管数量较多,粗细不一,直径范围为10~90 μm,平均直径约为70 μm(图2B、图2C、图2D),腺体小管管壁为单层柱状上皮细胞排列围成,且细胞核较大,但不同小管上皮细胞数量差异较大,有的较为集中,有的较为零散。在腺体小管的内腔表面可见微绒毛层,小管管腔内含有大量的分泌物质(图2B、图2C、图2D),且管腔内物质差异较大,有的较为致密,有的略稀疏,有的内容物较多,几乎填满整个管腔,有的则较少。腺体小管之间由致密结缔组织连接,结缔组织部分区域可见大量集中分布的血管和肌肉纤维,血管多为双层上皮细胞构成,肌肉纤维为平滑肌(图2E、图2F)。
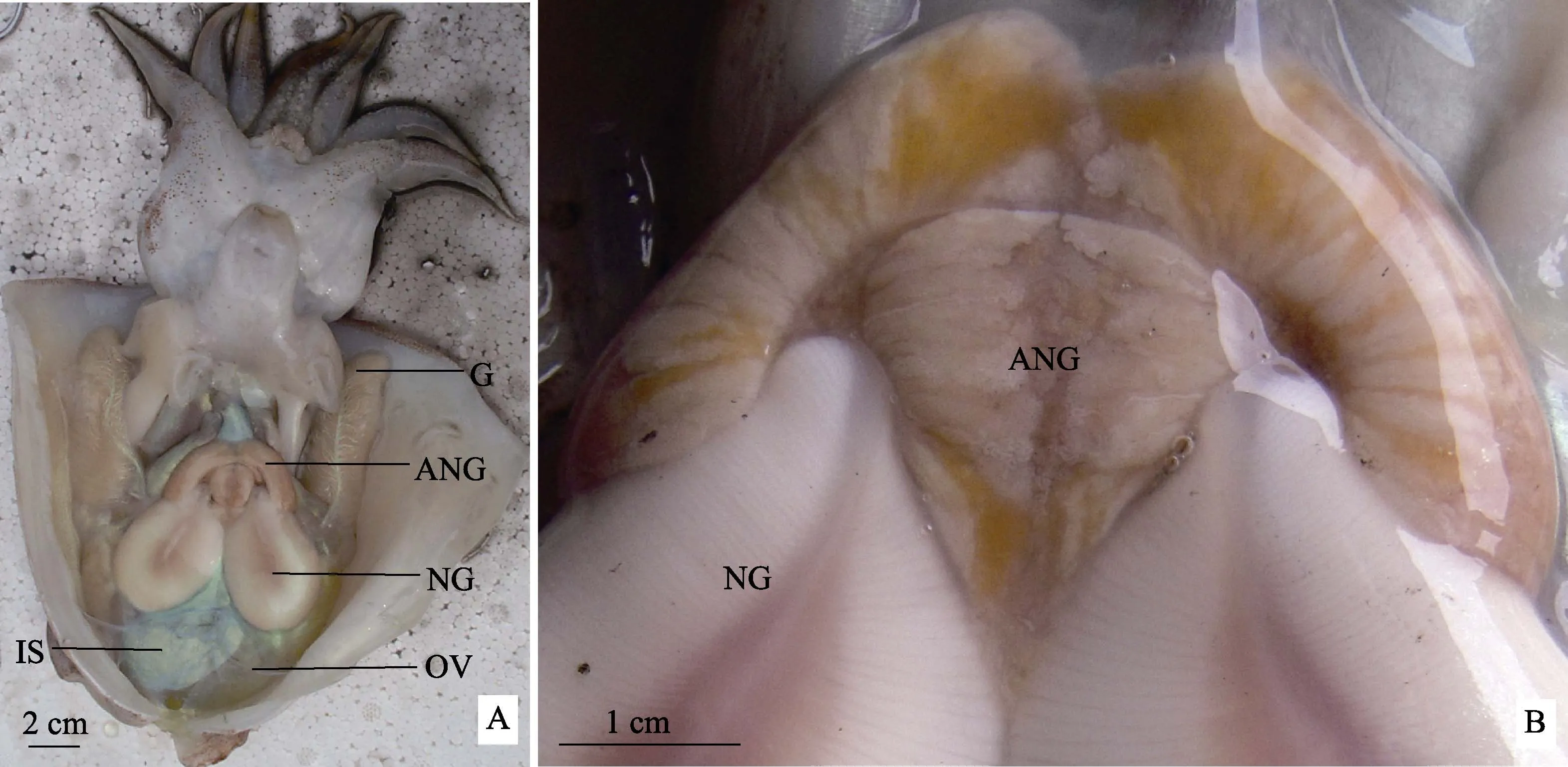
图1 金乌贼副缠卵腺的解剖结构Fig.1 Anatomical structure of accessory nidamental gland of S. esculentaA. 金乌贼内部构造;B. 副缠卵腺放大OV:卵巢;IS:墨囊;NG:缠卵腺;ANG:副缠卵腺;G:鳃A. The internal structure of S. esculenta; B. Enlarged accessory nidamental glandOV: Ovary; IS:Ink sac; NG: Nidamental gland; ANG: Accessory nidamental gland; G: Gill
![]()
图2 金乌贼副缠卵腺的显微结构Fig.2 Microstructure of accessory nidamental gland of S. esculentaA: 副缠卵腺横切面;B: 副缠卵腺横切面;C: 副缠卵腺横切面;D: 腺体管道横切面;E: 毛细血管横切面;F: 腺体管道横切面Mv:微绒毛;ML:肌肉层;OL:外膜层;Tu:腺体管道;Lum:管内腔;EC:上皮细胞;BV:血管;CT:结缔组织A: Transverse section of the accessory nidamental gland; B: Transverse section of the accessory nidamental gland;C: Transverse section of the accessory nidamental gland; D: Transverse section of the tubules;E: Transverse section of the blood vessels; F: Transverse section of the tubules Mv: Microvilli; ML: Musclar layer; OL: Outer layer; Tu: Tubules; Lum: Lumen; EC: Epithelial cells;BV: Blood vessel; CT: Connective tissue
2.3 副缠卵腺的超微结构
在金乌贼副缠卵腺的超微结构中,除管腔空白区域外,可见分布广泛的微绒毛区域、少量集中分布的纤毛区域以及数量较多的粗面内质网和囊泡(图3A)。2.3.1 小管上皮细胞 上皮细胞的细胞核较小,稍弯曲,呈蚕豆状,凸的一面称其为背部,凹的一面为腹部(图3B、图3C),在细胞质中有大量的线粒体,呈圆形或扁长形,且线粒体嵴的数量较多(图 3B、图3E、图3F),在细胞质内还存在大量集中分布的粗面内质网(图 3C、图 3F)和少量滑面内质网(图 3H)。细胞的基底膜平滑,由细胞外基质与结缔组织相连。在细胞质中圆形囊泡数量较多,且分布较为集中,部分囊泡中含有分泌球状体,且有些分泌球状体已释放到囊泡的外面,即腺体细胞内表面和管腔中(图3F、图3G、图3H、图3K)。在上皮细胞靠近管腔表面可见分布面积较广、密集排列的微绒毛,长度为2~4 μm,直径约为125 nm (图3A、图3D、图3G、图3H),此外,在微绒毛区域或靠近微绒毛区域可见集中分布(图 3G)或单行排列的纤毛(图 3H),纤毛与微绒毛方向平行或垂直,纤毛直径约 250 nm,轴丝具有典型的‘9+2’结构(图 3I)。
![]()
图3 金乌贼副缠卵腺的超微结构Fig.3 Ultrastructure of accessory nidamental gland of S. esculentaA: 副缠卵腺管道横切面;B: 小管上皮细胞;C: 上皮细胞放大;D: 小管上皮细胞;E: 上皮细胞细胞质;F: 上皮细胞细胞质;G: 小管上皮细胞近腔表面;H: 小管上皮细胞近腔表面;I: 纤毛放大;J: 腺体小管管腔;K: 管腔放大;L: 细菌放大Ve:囊泡;CI:纤毛;RER:粗面内质网;Mv:微绒毛;Lum:管内腔;N:细胞核;EM:细胞外基质;MI:线粒体;SG:分泌球状体;Ba:细菌;SER:滑面内质网;CM:中心微管;DM:双联体微管A: Transverse section of the accessory nidamental gland; B: Tubular epithelial cells; C: Amplification of the epithelial cells;D: Tubular epithelial cells; E: The cytoplasm of epithelial cells; F: The cytoplasm of epithelial cells; G: Near-cavity-surface ofthe tubular epithelial cells; H: Near-cavity-surface of the tubular epithelial cells; I: Amplification of the cilia;J: Lumen of the tubules; K: Amplification of the lumen; L: Amplification of the bacteria Ve: Vesicles; CI: Cilia; RER: Rough endoplasmic reticulum; Mv: Microvilli; Lum: Lumen; N: Nucleus;EM: Extracellular matrix; MI: Mitochondria; SG: Secretion globule; Ba: Bacteria; SER: Smooth endoplasmic reticulum;CM: Central microtubule; DM: Doublet microtubules
2.3.2 小管管腔 副缠卵腺的腺体小管管腔内具有大量的细菌,几乎占据整个管腔(图 3J、图 3K),但细菌的种类相对单一,绝大部分为具有双层膜的球菌(图3L),且未见正在分裂的细菌。此外,在管腔微绒毛处或靠近微绒毛处可见大量的上皮细胞产生的分泌球状体,球状体直径为0.1~0.4 μm,比细菌略偏小(图 3K)。
3 讨论
3.1 关于副缠卵腺的解剖结构
头足纲是软体动物门的重要经济种类,是重要的渔业对象,分为2个亚纲、8个目、6个亚目、46个科、13个亚科、147个属,共756种(陈新军等, 2009)。其中,有些种类具有副缠卵腺,如曼氏无针乌贼(蒋霞敏等, 2008)、拟目乌贼(Sepia lycidas)(罗江等,2014)、中国枪乌贼(Uroteuthis chinensis)(欧端木,1983)、莱氏拟乌贼(Sepioteuthis lessoniana)(Sivashanthini et al, 2010)、皮氏枪乌贼(Bloodgood,1977)等乌贼目和闭眼亚目(Myopsida orbigny)种类;而有些种类则不具有副缠卵腺,如真蛸(Octopus vulgaris)(Anna et al, 2001)、嘉庚蛸(Octopus tankahkeei)(许著廷等, 2011)、长蛸(Octopus variabilis)(许星鸿等, 2008)等蛸类,以及珍珠鹦鹉螺(Nautilus pompilius)(Have, 1977)等鹦鹉螺类属种。同乌贼目种类一致,金乌贼也具副缠卵腺1对且较发达,下端呈‘M’型与缠卵腺紧密连接,但副缠卵腺颜色存在一定差异,金乌贼产卵盛期颜色为白色并夹杂着橙黄色,而虎斑乌贼副缠卵腺在性成熟时由无色变为斑驳的红色(Nair et al, 2005)、曼氏无针乌贼性成熟时由白色变为橙黄色(王春琳等, 2010);皮氏枪乌贼的副缠卵腺在性成熟时体积增大,且颜色由白色变为亮红色(Bloodgood, 1977);欧洲横纹乌贼(Sepia officinalis)等头足类随着个体的发育,副缠卵腺由最初的白色变为黄色、橙色,性成熟后变为珊瑚红色,产卵后则变为粉红色(Richard et al, 1979),副缠卵腺颜色的差异原因可能与物间差异、管腔共生细菌产生的色素种类不同有关(王春琳等, 2010),也可能与一种类胡萝卜素Sepiaxanthin有关(Decleir et al, 1972),此外,王春琳等(2010)认为,副缠卵腺颜色变化可作为乌贼性成熟的“信号灯”。
3.2 关于副缠卵腺显微结构
同拟目乌贼(罗江等, 2014)、曼氏无针乌贼(王春琳等,2010)和皮氏枪乌贼(Bloodgood, 1977)的副缠卵腺显微结构一致,金乌贼副缠卵腺也主要为腺壁、腺体小管和结缔组织3部分构成,但腺壁的厚度存在一定差异,可能与品种差异或所处的发育阶段不同所致。此外,金乌贼副缠卵腺中管道数量也较多,且具有发达的结缔组织和毛细血管,因此,也是一种“管道”状的起分泌作用的腺体,其中,具分泌功能的是小管上皮细胞(王春琳等, 2010)。金乌贼副缠卵腺腺体小管管壁均为单层柱状上皮细胞排列围成,细胞呈扁平状,但福氏枪乌贼副缠卵腺中直径较小的小管由扁平细胞构成,直径略大的小管包含扁平细胞、纺锤状细胞和柱状上皮细胞3种细胞,而直径最大的小管仅存在柱状上皮细胞(Lum-Kong, 1992a),此外,Lum-Kong(1992a)研究还发现,未成熟的福氏枪乌贼雌性缠卵腺的腺体小管较细,且内部中空,性成熟后,腺体小管数量增多,充满细菌和分泌物。金乌贼副缠卵腺小管较多,且管腔较粗,管内物质差异较大,有的较为致密,有的略稀疏,有的内容物较多,有的则较少,由于金乌贼处于产卵盛期,其原因可能与产卵损耗所致,但与福氏枪乌贼不同(Lum-Kong, 1992a),金乌贼副缠卵腺的管腔内物质未见浅色区域,因此,不同头足类种类其显微结构存在一定差异。
3.3 关于副缠卵腺超微结构
金乌贼副缠卵腺超微结构研究表明,副缠卵腺是一种起分泌作用的腺体,主要体现在:(1)在上皮细胞的细胞质囊泡中、内表面、微绒毛丛和管腔中含有较多呈颗粒状散布的分泌球状体,说明了球状体在上皮细胞中形成,并逐渐向细胞内表面迁移,直至从管腔排出的过程,这与曼氏无针乌贼(王春琳等, 2010)、枪乌贼(Loligo opalescens)(Kaufman et al, 1998)和虎斑乌贼(Nair et al, 2005)等副缠卵腺中的球状体观察结果一致;(2)在上皮细胞靠近管腔表面可见大量的微绒毛及少量的纤毛,二者通过协助运动将管腔内的分泌物及共生菌等运送出管道外。但头足类不同种类间微绒毛和纤毛的排列方式存在一定差异,如金乌贼微绒毛密集排列,分布连续,纤毛只分布在微绒毛区域或靠近微绒毛区域,分布集中或呈单行排列,着生不连续,这与曼氏无针乌贼(王春琳等, 2010)、虎斑乌贼(Nair et al, 2005)基本相同,但皮氏枪乌贼微绒毛和纤毛均着生连续(Bloodgood, 1977)。林东明等(2013)研究表明,头足类副缠卵腺的分泌物为凝胶状物质,可在乌贼产卵时参与三级卵膜的形成,此外,其分泌物还具有黏连卵串的作用(王津伟等, 2010)。同曼氏无针乌贼(王春琳等, 2010)、乌贼(Sepia officinalis)(Branden et al, 1980; Grigioni et al, 2000)、皮氏枪乌贼(Bloodgood, 1977)、福氏枪乌贼(Lum-Kong, 1992a)、微鳍乌贼科(Idiosepiidae)(Pichon et al, 2005)等头足类一致,金乌贼副缠卵腺的腺体小管腔内也充满着大量共生细菌。但不同的头足类共生菌的种类存在一定差异,如曼氏无针乌贼副缠卵腺中细菌呈棒状或杆状,且有正在分裂的细菌(王春琳等, 2010),虎斑乌贼(Nair et al, 2005)和皮氏枪乌贼(Bloodgood, 1977)副缠卵腺中细菌为球菌和杆菌。Grigioni等(2000)鉴定出乌贼的副缠卵腺中细菌主要分为 Agrobacterium,Roseobacter, Sporichthya, Rhodobium-Xanthobacter和Clostridium共5个属。金乌贼副缠卵腺中的细菌种类相对单一,绝大部分为球菌,且未见正在分裂的细菌,其共生菌的种类有待进一步分离鉴定。
由于头足类副缠卵腺在孵化后一段时间才开始出现,且开口于外套腔和海水中,因此,王春琳等(2010)和Kaufman等(1998)研究表明,共生菌是在副缠卵腺的小管形成后,在乌贼吸水的过程中进入,属于“水平传递”,而不是Buchner等(1965)认为的来自于上一代亲体副缠卵腺的“垂直传递”。Grigioni等(2000)和Barbieri等(1997)研究表明,这些共生菌可产生抗生素或细菌毒素等抑菌物质,但Nair等(2005)研究表明,只有在乌贼最终成熟后才能产生抗菌活性。乌贼受精卵具有三级卵膜结构,其中,第3层卵膜是卵子释放到外套腔时,被缠卵腺和副缠卵腺分泌物包裹所形成(Boletzky, 1986),因此,在受精卵的卵膜中含有一定数量的共生菌。由于金乌贼受精卵孵化水温较高且孵化时间较长(近30 d),挂卵附着基上受精卵集中区域极容易发生腐烂,共生菌的存在可在一定程度上保护受精卵免受病原体的侵害(Grigioni et al, 2000),进而提高孵化率,因此,副缠卵腺中含有共生菌可能是金乌贼的繁殖策略之一。此外,关于共生菌的其他作用,Boucher-Rodoni等(1994)研究表明,这些细菌可驱动体内氨基态的氮转化为氮气,然后进入内壳而产生浮力。目前,关于金乌贼副缠卵腺共生菌的研究资料空白,其共生菌来源以及抗菌和浮力调节作用等有待进行深入研究。
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!
