
论文:豆粕替代鱼粉对牛蛙生长性能、消化酶活性和肠道健康的影响
豆粕替代鱼粉对牛蛙生长性能、消化酶活性和肠道健康的影响
丁 李,王 玲,鲁康乐,宋 凯,张春晓
(集美大学水产学院,厦门市饲料检测与安全评价重点实验室,福建厦门 361021)
随着水产养殖业迅速发展,鱼粉作为水产养殖动物饲料的主要蛋白源,其需求量急剧上升[1],然而由于世界渔业资源的逐年衰退,导致鱼粉的价格居高不下,因此寻找可替代鱼粉的优质的蛋白源成为水产养殖业可持续发展的重要课题之一[2-4]。豆粕含有较高的可消化蛋白质,价格低廉,成为替代鱼粉的主要植物蛋白源[5]。但是豆粕中含有大量的抗营养因子如蛋白酶抑制剂、凝集素、植酸和抗维生素以及难以消化的碳水化合物等,长期大量食用会对鱼、虾等水产养殖动物肠道健康造成负面的影响[4、6-7]。因此,有必要研究植物蛋白对养殖动物生理生化的影响。
牛蛙(Ranacatesbeiana)原产于北美洲,在分类学上属于脊索动物门两栖纲无尾目蛙科蛙属,是大型杂食性蛙类[8],于1959年从古巴引入我国。因其具有生长速度快,适应能力强、肉质鲜美、产量高等优点,近年来,牛蛙养殖业在我国南方地区发展迅速,成为新兴的特种水产养殖品种之一[9]。鱼粉是牛蛙饲料的重要蛋白源,本实验室前期研究表明,豆粕可替代80%的鱼粉而不影响其生长性能,但当豆粕100%替代鱼粉时会显著降低其生长性能[10]。在此基础上本实验进一步研究豆粕部分或完全替代鱼粉对牛蛙生长性能、肠道健康、消化酶活性和饲料消化率的影响,以期为牛蛙饲料中植物蛋白的大量使用提供理论基础。
1 材料与方法
1.1 试验饲料
以鱼粉为蛋白源,鱼油和豆油为主要脂肪源配制基础饲料,基础饲料中鱼粉的添加水平为44%,以基础饲料为对照,以豆粕分别代替基础饲料中50%和100%的鱼粉,并添加三氧化二钇作为内源指示剂,配制3组等氮等能的试验饲料,分别记为FM、B1和B2,试验饲料的原料组成和营养成分见表1。各饲料原料分别粉碎,过60目筛网,所有原料按照配方等比例逐级依次混合,使用水产饲料膨化机制成膨化饲料,20 ℃条件下自然风干至水分约10%,然后保存于-20 ℃冰箱中备用。
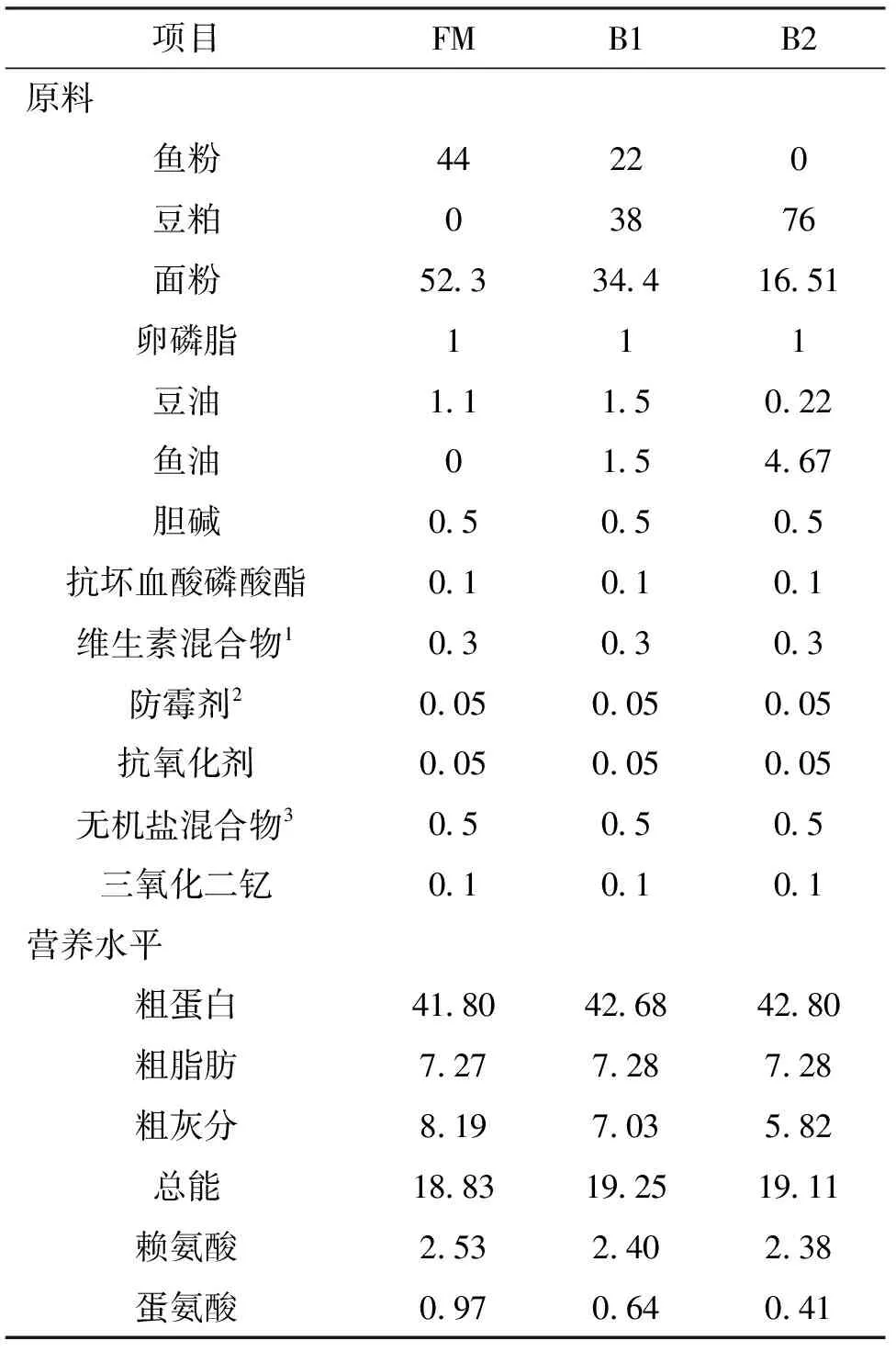
表1 试验饲料配方和营养组成
注:1.每千克维生素混合物含VB110 mg,核黄素8 mg,盐酸吡哆醇10 mg,VB120.2 mg,VK310 mg,肌醇100 mg,泛酸钙20 mg,烟酸50 mg,叶酸2 mg,生物素2 mg,VA(50万IU)400 mg,VD 5 mg,VE(50万IU)100 mg,乙氧基喹啉150 mg,次粉0.1328 g。2.50%丙酸钙和50%富马酸。3.每千克矿物质混合物含氯化钾200 mg,碘化钾60 mg,硫酸钴(1%)100 mg,硫酸铜24 mg,硫酸亚铁400 mg,硫酸锌174 mg,硫酸锰78 mg,硫酸镁800 mg,亚硒酸钠(1%)50 mg,沸石粉3.114 g。
1.2 养殖试验管理
牛蛙购于厦门市翔安区某养殖场,为同批次培育的蛙苗。实验开始前将牛蛙暂养2周,期间投喂基础饲料。暂养结束后,停饲24 h,挑选大小均匀并且健康的牛蛙[初始平均体重为(40.0±0.3)g]随机分配到15个玻璃缸内(容积为0.7 m×0.5 m×0.4 m)。试验随机分为3个组,每组5个重复,每个重复放10只牛蛙。在试验期间,水温为(26±3)℃,水位在5 cm左右,养殖周期为30 d。每天饱食投喂2次(8:00和18:00),投喂前1 h清理玻璃缸并换水。
1.3 样品采集
养殖实验结束后,停饲24 h,分别对每缸牛蛙计数并称重;每个饲料组随机选取3个玻璃缸进行采样,剩下的2个玻璃缸继续养殖,用于饲料消化率研究。每缸分别取9只牛蛙,用双毁髓法处死。每只牛蛙解剖后取空肠和回肠,即牛蛙十二指肠和直肠之间的部分,肠壁较厚的前半部分为空肠,肠壁较薄的后半部分为回肠,分别装入灭菌的冻存管中,迅速放入液氮保存,其中3只牛蛙用于消化酶活性的测定,3只用于相关炎性因子基因表达的测定。剩余3只牛蛙的空肠和回肠各取1 cm放入伯恩溶液中固定,用于肠道切片制作。粪便的收集参考文献[11]的方法,收集包膜完整的粪便放入-20 ℃的冰箱中保存。
1.4 试验指标测定
1.4.1 常规指标分析
饲料和粪便的粗蛋白、粗脂肪、粗灰分、水分的含量测定参考AOAC(2005)的方法进行,粗蛋白采用杜马斯燃烧定氮法以全自动蛋白质分析仪(Elementer rapid N EXCEDD)测定,粗脂肪采用的是索氏抽提法测定,粗灰分采用马弗炉550 ℃高温灼烧法测定,水分采用105 ℃恒重法测定;
1.4.2 消化酶测定
淀粉酶、脂肪酶和总蛋白试剂盒购于南京建成生物研究所,蛋白酶试剂盒购于上海盈公生物科技有限公司;饲料和粪便中钇采用电感耦合等离子发射光谱仪(ICP-OES Prodigy7,Leeman,USA)进行测定;饲料和粪便的能量采用全自动氧弹量热仪(Parr 6300)测定。
1.4.3 肠道组织观察
从伯恩固定液中分别取出空肠和回肠组织各0.5 cm,用生物组织自动脱水机(KD-TS3A)进行不同浓度酒精脱水,石蜡包埋组织,修整凝固蜡块,用组织切片机(型号:德国leica公司生产的RM2235切片机)切片,厚度为6 μm;脱蜡后用苏木精-伊红(H.E)染色,封片后用正置荧光显微镜(Leica DM5500B)观察测定和拍照。
1.4.4 肠道炎性因子测定
使用Trizol试剂(TaKaRa)分别提取空肠和回肠组织的总RNA,使用TaKaRa公司的逆转录试剂盒将mRNA进行反转录;使用SYBR Green RT-PCR试剂盒(TaKaRa)在Thermal cycler (ABI StepOne PlusTM)Applied Biosystems QuantStudio 6&7 系统上进行实时荧光定量,分析肠道组织促炎性因子mRNA(引物序列见表2)表达量的差异。使用β-actin作为内参基因,2-ΔΔCt法计算各炎性因子基因的相对表达量。
![]()
表2 引物列表
1.5 计算公式
增重率(WG)=(Wf-Wi)/Wi×100%
特定生长率(SGR)=(lnWf-lnWi)×100%/t
饲料效率(FE)=(Wt-Wo)/Wd
摄食率(FR)=Wd/(Wt/2-Wo/2)/t×100%
蛋白质效率(PER)=(Wt-Wo)/Wp
成活率(SR)=Nf/Ni×100%
式中Wi和Wf分别表示牛蛙初始均重和末期均重,Wp为总蛋白摄入量,Wd为摄入饲料总重,Nf为末蛙数,Ni为初始蛙数,t为试验天数,Wt为牛蛙终末总重,Wo为牛蛙初始总重。
1.6 数据分析
实验数据处理采用SPASS 22.0软件进行单因素方差分析处理(one-way ANOVA),若差异显著,则进行Duncan氏法多重比较,P<0.05时差异显著,数据用平均值±标准差(Mean±SD)表示,柱形图使用Prism 5软件绘制。
2 结果
2.1 豆粕替代鱼粉对牛蛙生长性能和饲料利用率的影响
试验各组成活率均为100%。豆粕替代组(B1和B2)牛蛙的终末均重(FBW)、增重率、特定生长率、饲料效率、蛋白质效率显著低于FM;而FR在B1组显著高于FM和B2组,FM和B2组之间差异显著。
![]()
表3 豆粕替代鱼粉对牛蛙生长性能的影响
注:同行数据肩标不同小写字母表示差异显著(P<0.05),相同小写字母表示差异不显著(P>0.05)。下表同。
2.2 牛蛙对试验饲料营养物质的表观消化率
由表4所示,随着豆粕替代水平的增加,饲料干物质的表观消化率显著降低,变化范围为78%~86%;豆粕替代鱼粉后显著降低了牛蛙的蛋白质表观消化率,但B1和B2组间无显著差异;能量的表观消化率随替代水平的增加显著降低,变化范围为79%~86%。
![]()
表4 豆粕替代鱼粉对牛蛙表观消化率的影响
2.3 豆粕替代鱼粉对牛蛙肠道消化酶活性的影响
本实验中牛蛙空肠蛋白酶活性在B2组显著低于FM和B1组,FM和B1组间无显著差异;而脂肪酶和淀粉酶活性在各替代组之间没有显著差异;回肠蛋白酶活性也在B2组最低,其他两组间无显著差异;脂肪酶活性在FM组最高,显著高于B1和B2组;而淀粉酶活性则在FM组显著低于B1和B2组。
![]()
表5 豆粕替代鱼粉对牛蛙肠道消化酶的影响(n=3)
2.4 豆粕替代鱼粉对肠道形态学的影响
如表6所示,豆粕替代鱼粉显著影响牛蛙肠道绒毛高度、肌层厚度和柱状上皮高度。在空肠和回肠,随着豆粕替代水平的升高,绒毛高度、肌层厚度和柱状上皮高度都呈现显著降低的趋势。从图1和图2看出,FM组肠道绒毛完整,绒毛密集且较长,柱状上皮排列整齐,无上皮细胞自溶现象发生;随着豆粕替代水平的升高,肠壁变薄,肠绒毛变短且稀疏,出现上皮细胞自溶的现象,细胞核排列紊乱,柱状上皮变窄;特别在B2组上皮细胞自溶现象明显,细胞核出现聚集现象,肠绒毛变短且稀疏,柱状上皮排列不规则且变薄。
![]()
表6 豆粕替代鱼粉对牛蛙肠道组织结构的影响(n=3)
![]()
图1 豆粕替代鱼粉对牛蛙空肠组织结构的影响Fig.1 Effects of replacement fish meal by soybean meal on the tissue structure for the jejunum of R.castesbeiana1.FM;2.B1;3.B2; V.绒毛;M.肌层;CE.柱状上皮。下同。
![]()
图2 豆粕替代鱼粉对牛蛙回肠组织结构的影响Fig.2 Effects of replacement fish meal by soybean meal on the tissue structure for the ileum of R.castesbeiana
2.6 豆粕替代鱼粉对肠道炎性因子基因表达量的影响
图3显示,牛蛙空肠炎性因子基因IL-1β、IL-8和IL-17表达量在B2组最高,显著高于FM和B1组,但后两者之间没有显著差异;炎性因子TNF-α基因表达量随着豆粕替代水平的升高而显著升高。从图4可以看出回肠中IL-1β和IL-17基因的表达量随着豆粕替代水平的升高而显著升高,而IL-8和TNF-α基因表达量在B2组都显著高于FM组,但FM和B1组间差异不显著。
![]()
图3 豆粕替代鱼粉对牛蛙空肠炎性因子基因mRNA表达量影响Fig.3 Effects of replacement fish meal by soybean meal on mRNA expression of inflammatory factor gene for the jejunum of R.castesbeiana1.FM;2.B1;3.B2;数据柱形图中标注小写字母不同表示差异性显著(P<0.05)。下同
![]()
图4 豆粕替代鱼粉对牛蛙回肠炎性因子基因mRNA表达量影响Fig.4 Effects of replacement fish meal by soybean meal on mRNA expression of inflammatory factor gene for the ileum of R.castesbeiana
3 讨论
豆粕作为主要的植物蛋白源已经在水产动物饲料中广泛应用,但与鱼粉相比,豆粕的氨基酸并不平衡,并且含有大量抗营养因子,致使养殖动物摄食大量豆粕后生长性能和饲料利用率会受到负面影响[12-13]。本研究结果也发现,在不配平氨基酸的情况下,豆粕替代50%和100%鱼粉显著降低了牛蛙的生长性能。然而,方卫东等[10]的研究结果表明,补充赖氨酸和蛋氨酸后,豆粕替代80%的饲料鱼粉而不影响牛蛙的生长性能。在本实验中,随着豆粕替代水平的增加,饲料赖氨酸和蛋氨酸逐渐降低,赖氨酸和蛋氨酸是水产动物的必需氨基酸,对鱼类的研究表明,饲料赖氨酸和蛋氨酸缺乏会降低其生长性能、饲料利用率和蛋白质沉积率,增加死亡率[14]。上述研究结果说明氨基酸平衡是限制牛蛙饲料中豆粕替代鱼粉水平的重要因素。众多研究表明,豆粕替代水平升高会导致银色黑鲷(Sparidentexhasta)[13]等水产动物摄食率下降。但在本研究发现B1组的牛蛙摄食率最高,显著高于B2组和FM组,这与Zhang等[15]对花鲈(Lateolabraxjaponicus)和Taher等[16]对游泳蟹(Portunuspelagicus)的研究结果一致,都随着豆粕替代水平的升高,摄食率表现出先升高后降低的趋势。
营养物质的消化率是评价饲料营养价值的主要指标。一般来说,水产动物对动物蛋白饲料的消化率比对植物蛋白饲料要高[13]。本实验结果显示,豆粕替代鱼粉显著降低了牛蛙对饲料干物质、蛋白质和能量的表观消化率,这与对大西洋鳕鱼(Gadusmorhua)(24%)[17]、欧洲鲈(Dicentrarchuslabrax)(50%)[18]和高体鰤(Serioladumerili)[19]等的研究结果一致,可能是因为豆粕中含有的抗营养因子降低了饲料营养物质的表观消化率。另外饲料营养成分的消化率与动物消化道内消化酶的活性有关[20],同时消化酶活性也受饲料组成的影响[21-22]。已有研究表明,豆粕高比例替代鱼粉会降低罗非鱼(Oreochromisniloticus×O.aureus)[23]、牛蛙[10]和花鲈[15]等养殖动物肠道消化酶活性。本试验也发现,豆粕100%替代鱼粉显著降低了牛蛙肠道蛋白酶活性以及回肠脂肪酶活性,其变化趋势与表观消化率结果相似,说明饲料中高水平的豆粕会影响牛蛙肠道消化酶的活性,从而降低牛蛙对饲料的表观消化率。
肠道是水产动物对营养物质消化吸收的主要器官。本试验中,豆粕替代组牛蛙肠绒毛高度,肌层厚度以及柱状上皮高度都出现降低现象,表明豆粕替代鱼粉可能改变了牛蛙肠道组织结构,在大黄鱼[24]、建鲤(CyprinuscarpiovarJian)[25]、高体鰤[19]和花鲈[15]的研究中也有相似现象,这可能是由于肠上皮细胞表面的特异性受体与豆粕中的某些抗营养因子结合,导致肠黏膜出现炎症反应所致。已有研究表明,豆粕抗营养因子凝集素与多糖可能会破坏肠道微绒毛结构,减少了营养物质的吸收[26],而大豆抗原蛋白可以引起鱼类肠道炎症,从而可能影响肠道结构,对肠道健康产生负面影响[27]。在本研究中,随着豆粕替代水平的增加,牛蛙肠道促炎症因子TNF-α,IL-1β,IL-8和IL-17表达量显著增强。在很多鱼类研究中也发现了类似现象[28-30],这说明豆粕可能通过诱导牛蛙肠道炎症反应,进而导致肠道组织结构改变。
4 结论
在本实验条件下,50%豆粕替代鱼粉即会降低牛蛙的生长性能、饲料利用率及其对饲料营养物质的表观消化率;高豆粕饲喂牛蛙可诱导其肠道炎症反应,豆粕诱导的肠道炎性反应可能是损害肠道健康、降低营养物质消化吸收的重要因素。
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!
